
ARTÍCULO ESPECIAL
JONNPR y las investigaciones realizadas en el camino al Premio Nobel 2019. Una visión personal sobre las moléculas y los aspectos moleculares y mecanismos de control subyacentes relacionados con la hipoxia y el cáncer
JONNPR and the research carried out on the way to the Nobel Prize 2019. A personal view on the molecules and the molecular aspects and underlying control mechanisms related to hypoxia and cancer
Francisco J. Sánchez-Muniz1, Jesús M Culebras2, Luis Vicente-Vacas3
1 Catedrático de Nutrición. Facultad de Farmacia de la Universidad Complutense de Madrid y Académico de número de la Real Academia Nacional de Farmacia, España
2 De la Real Academia de Medicina y Cirugía de Valladolid y del IBIOMED, Universidad de León. Director de Journal of Negative & No Positive Results. España
3 Editor de Journal of Negative & No Positive Results. España
* Autor para correspondencia.
![]()
|
|
Attribution-NonCommercial-ShareAlike 4.0 International License La revista no cobra tasas por el envío de trabajos, |
|
Resumen
Este artículo especial desgrana los mecanismos y aspectos centrales relacionados con tres moléculas: la eritropoyetina, el factor inducible por hipoxia (HIF) y la proteína von Hippel-Lindau (pVHL) que han dado lugar tras múltiples investigaciones a galardonar con el Peremio Nobel de Fisiología y Medicina 2019 a los científicos William G. Kaelin, Gregg L. Semenza y Peter J. Ratcliffe. Se señalan los mecanismos moleculares que tiene lugar en situación de normoxia, hipoxia y pseudohipoxia. El artículo termina señalando las perspectivas farmacológicas que ha abierto la relación investigadora de estos científicos en el campo del cáncer y de muchas enfermedades degenerativas.
Palabras clave
Premios Nobel, hipoxia, normoxia, HIF, VHL, nuevas perspectivas farmacológicas
Abstract
This special article discusses the mechanisms and central aspects related to three molecules: erythropoietin, hypoxia-induced factor (HIF) and von Hippel-Lindau protein (pVHL) that have led after multiple investigations to win the Nobel Prize in Physiology and Medicine 2019 to scientists William G. Kaelin, Gregg L. Semenza and Peter J. Ratcliffe. The molecular mechanisms taking place in a situation of normoxia, hypoxia and pseudohypoxia are indicated. The article ends by pointing out the pharmacological perspectives that the research of these scientists has opened in the field of cancer and many degenerative diseases.
Keywords
Nobel Prize, hypoxia, normoxia, HIF, VHL, pharmacological perspectives
En nuestro homenaje a Alfred Nobel y a los galardonados con el premio Nobel de Fisiología y Medicina de 2019(1) señalábamos que según la Fundación Nobel(2) la investigación de estos premiados tiene como protagonistas principales a tres moléculas: la hormona eritropoyetina (EPO), el Factor inducible por hipoxia (HIF) y la proteína Von Hippel-Lindau (pVHL). En este artículo especial comentaremos con detalle algunos aspectos moleculares y los mecanismos subyacentes relacionados con la hipoxia y los estados pseudohipóxicos del cáncer y otras patologías.
a) La vía metabólica de la hipoxia y su relación con la investigación de los premiados. El grupo de investigación del genetista pediátrico Semenza, investigaba sobre los mecanismos desencadenantes de la producción de eritropoyetina, un factor de crecimiento hematopoyético producido por el hígado y los riñones que promueve la generación de glóbulos rojos(3).

Greeg L Semenza.
Galardonado con el Premio Nobel de Medicina 2019
Wang y Semenza(4) identificaron un sitio de unión específico a la secuencia del ADN (denominado elemento de respuesta a la hipoxia [HRE]) para un factor de transcripción en la región anexa al extremo 3' del gen de la eritropoyetina humana (EPO) y posteriormente purificaron un factor de transcripción, al que Semenza llamó factor inducible por hipoxia 1 (HIF-1)(4).
El HIF se halla presente en todas las células de mamíferos estudiadas y, remontando en la escala evolutiva, se le encuentra hasta en el nemátodo C. elegans y la mosca drosofila D. melanogaster(5). El complejo HIF contiene una subunidad reguladora alfa y una subunidad constitutiva beta, que pertenecen a la familia de los factores de transcripción bHLH/PAS (proteínas que contienen además del dominio bHLH (por basic hélix-loop-helix)(5), un dominio PAS (por Per Arnt Sim).
La subunidad beta es constitutiva y está asimismo implicada en la respuesta a los xenobióticos que se acompaña de una heterodimerización con el receptor intracelular AhR (Aryl hydrocarbon Receptor). La subunidad alfa es regulada por el oxígeno. Hasta la fecha han sido identificados en los mamíferos tres genes distintos, que codifican la subunidad HIF-α: HIF-1α, HIF-2α (también llamado EPAS1) y HIF-3α (uno de cuyos transcritos ha sido llamado IPAS, por inhibitory PAS protein (proteína inhibidora a dominio PAS)(5). Como regla general, los cultivos celulares expresan ambos genes HIF-1α e HIF-2α, aun cuando no está todavía completamente dilucidada la importancia de cada uno de ellos. Los dos factores inducibles por la hipoxia, HIF-1 y HIF-2 son los principales mediadores de la adaptación celular a la hipoxia y se hallan fuertemente expresados, en condiciones de hipoxia tanto regional como sistémica. HIF-2 parece estar más en relación con la secreción de EPO por los fibroblastos maduros intersticiales(5).
Al producirse un estado de hipoxia sistémica, como es el caso de la anemia, se induce la formación de HIF-1 y 2. Sin embargo, señalaremos que por ejemplo, las células tubulares renales expresan sólo HIF-1, mientras que el HIF-2 se encuentra en las células glomerulares, las células endoteliales peritubulares y los fibroblastos intersticiales. Es decir, HIF-1 y HIF-2 parecen ejercer funciones complementarias in vivo(5).
También se ha definido la existencia de un alelo “especial” del gen HIF responsable de la adaptación a grandes altitudes en humanos(6,7). En altitudes elevadas, la proteína EPAS1 o HIF-2a es útil como respuesta adaptativa a corto plazo. Sin embargo, EPAS1 puede originar una producción excesiva y crónica de eritrocitos, lo cual puede causar la muerte y la inhibición de las capacidades reproductivas. No obstante, hay evidencia científica de que ciertas variantes del gen de la HIF proporcionan protección para aquellos habitantes que viven a una altura sobre el nivel del mar próxima a 4000 metros, donde las concentraciones de oxígeno atmosférico están ampliamente reducidas (aprox. un 40%).
La proteína codificada contiene un dominio de proteína hélice-bucle-hélice básica, así como otro dominio identificado en las proteínas involucradas en rutas de transducción de señales que responden a los niveles de oxígeno del ambiente(5,8,9,10). El gen HIF (componente 2a) está implicado en el desarrollo del corazón embrionario y se expresa en las células endoteliales de las paredes de los vasos sanguíneos del cordón umbilical para mantener el control homeostático de las catecolaminas y con ello prevenir el riesgo de insuficiencia cardíaca(9). El dímero HIF, además de inducir la expresión génica de la eritropoyetina, induce a otros genes que son muy necesarios para una respuesta correcta celular y sistémica frente a la hipoxia y el mantenimiento de la homeostasis celular (Figura 1). Entre ellos estaba el gen del factor de crecimiento endotelial vascular (en inglés VEGF, de vascular endotelial growth factor gene), que desempeña papeles importantes en la angiogénesis(11).
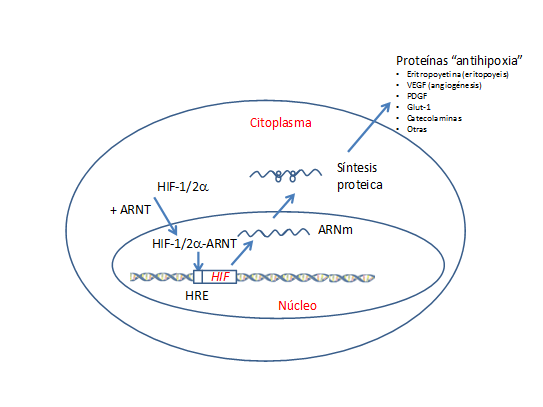
Figura 1. Situación de hipoxia. La falta de disponibilidad de oxígeno conduce a una mayor presencia del factor de transcripción inducible por hipoxia alfa (HIF1/2a), que se transloca al núcleo uniéndose al translocador nuclear del receptor de aril-hidrocarburo (ARNT). En el núcleo se une a la secuencia más próxima del gen HIF en una zona conocida como elemento del receptor del gen HIF-1/2 o HRE, poniendo en marcha la maquinaria de expresión génica con la transcripción de ARN mensajeros y su traducción posterior en los ribosomas para formar múltiples proteínas relacionadas con mecanismos protectores contra la hipoxia, promoción de la vascularización (angiogénesis), utilización metabólica, etc. (Esquema de diseño propio)
b) La molécula VHL y su conexión con el cáncer. Simultáneamente a los estudios de Semenza, el grupo de Kaelin estaba trabajando en la enfermedad VHL, una enfermedad autosómica dominante con afectación multisistémica(12) que incluye dolores de cabeza, sensación de vértigo, marcha inestable, mareos, debilidad de las extremidades, problemas de visión y de presión arterial. Asociadas a dicha enfermedad aparecen angiomatosis, hemangioblastomas, feocromocitoma, carcinoma de células renales, quistes pancreáticos (cistadenoma seroso pancreático), tumor del saco endolinfático y cistadenomas papilares bilaterales del epidídimo o ligamento ancho del útero(13,14).

Williams G. Kaelin
Galardonado con el Premio Nobel de Medicina 2019
La enfermedad es causada por mutaciones del gen supresor tumoral de von Hippel-Lindau (VHL) en el brazo corto del cromosoma 3 (3p25-26)(15). Hay más de 1500 mutaciones en la línea germinal y mutaciones somáticas en la enfermedad de VHL(16). El 30-40% de las mutaciones en el gen VHL consisten en mutaciones de deleción de 50-250 kb que eliminan parte del gen o todo el gen y regiones vecinas del ADN. El 60-70% restante de la enfermedad VHL es causado por el truncamiento de pVHL por mutaciones “sin sentido”, o mutaciones que atañen a la unión de los exones(16).
Kaelin, formado como internista (fue residente en jefe del servicio Johns Hopkins Osler) y oncólogo, estaba intrigado por el hecho de que los tumores asociados al síndrome de VHL siempre eran de naturaleza muy vascular y porque los pacientes afectados ocasionalmente presentaban policitemia secundaria. Estudios bioquímicos específicos indicaron que la proteína del gen VHL (pVHL) forma un complejo con papel decisivo en la degradación de proteínas específicas tras su ubiquitinación (Figura 2).
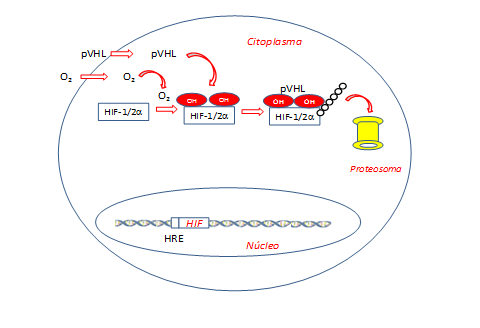
Figura 2. La presencia de oxígeno (normoxia) induce la activación del sistema prolil-hidroxilasa e incrementa la presencia de la proteína VHL (pVHL). Esta proteína tiene actividad ubiquitina ligasa y es responsable de añadir restos ubiquitina a la proteína HIF hidroxilada, permitiendo el reconocimiento por el proteosoma y su degradación posterior, evitando que HIF pueda inducir la formación de proteínas anti-hipoxia. HIF: Factor inducible por hipoxia. (Esquema de diseño propio)
c) La proteína VHL y la hipoxia. Regulación de la interacción VHL-HIF. Una observación crucial que conectó las líneas de investigación del grupo de Semenza con el de Kaelin provino del grupo de Piter J Ratcliffe, un nefrólogo muy intrigado en conocer cómo los riñones servían como órganos sensores de oxígeno y, por lo tanto, capaces de regular por vía sistémica la producción de eritropoyetina.
En los primeros experimentos, el grupo de Ratcliffe demostró que múltiples tipos de células eran capaces de detectar la hipoxia y conducir la transcripción de EPO a través del HRE que Semenza había identificado(18). Esto sugirió que HIF posiblemente servía como un mecanismo de respuesta universal a la hipoxia. Es importante destacar que, en 1999, el equipo de Ratcliffe demostró que pVHL regulaba la proteína HIF-1α, actuando probablemente como su ubiquitina ligasa E3(19). Por tanto Semeza y Ratcliffe descubrieron de forma independiente que el “sensor” de oxígeno estaba presente en diferentes células corporales y compartían un mecanismo común de detección de oxígeno.

Sir Piter J Ratcliffe
Galardonado con el Premio Nobel de Medicina 2019
El mecanismo por el que se producía la estabilización o la activación del programa transcripcional masivo en respuesta a la hipoxia se delimitó por los tres grupos mediante un modelo relativamente sencillo. Así, pVHL está involucrada en la regulación de la proteína HIF-1α. Como hemos previamente comentado, HIF-1α es una subunidad del factor de transcripción heterodimérico HIF que a niveles normales de oxígeno celular está altamente regulado. En condiciones fisiológicas normales, pVHL reconoce y se une a HIF-1α solo cuando hay oxígeno debido a la hidroxilación postraduccional de dos residuos de prolina en la proteína HIF-1α (Figura 2).
El proceso delimitado en la Figura 2 sugirió que la regulación de HIF es postranscripcional: HIF-1α se transcribe, traduce y degrada rápidamente, a menos que el oxígeno se vuelva limitante. Así, se identificó una segunda proteína homóloga, HIF-2α, que presenta una regulación dependiente de hidroxilación idéntica a través de pVHL. Cuando se estabiliza el HIF-1/2α, se heterodimeriza rápidamente con HIF-1β (no presentado en los esquemas), transcribiendo cientos de genes que son críticos para la respuesta celular a la hipoxia entre los que se incluyen el VEGF, el gen del factor de crecimiento derivado de plaquetas B (PGFB), gen de la eritropoyetina (EPO) y genes involucrados en la captación de glucosa y el metabolismo, de enorme importancia en la homeostasis y la supervivencia(20,21) (Figura 1).
En condiciones de baja concentración de oxígeno o en casos de enfermedad de VHL, donde el gen VHL está mutado (Figura 3), la proteína pVHL no se une a HIF-1α, ya que al no ser reconocida escapa de ser degradada(22,23). Como hemos comentado la pVHL presenta actividad ligasa tipo E3 que ubiquitina a HIF-1α y causa su degradación por el proteosoma (Figuras 2 y 3)
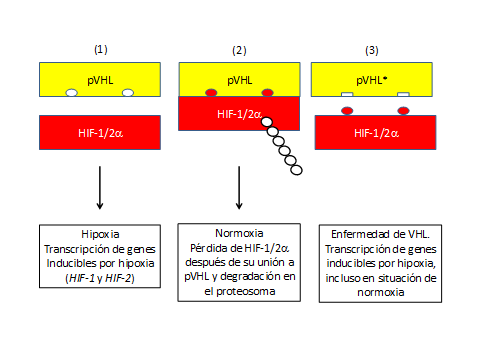
Figura 3. Regulación de HIF-1/2alfa por la proteína pVHL. (1) Hipoxia; (2) Normoxia con presencia adecuada de oxígeno o hiperoxia en el caso de exceso de oxígeno; (3) Enfermedad de von Hippel-Lindau (VHL). Se señala con dos círculos rojos la hidroxilación de HIF. En el caso de la enfermedad de von Hippel Lindau a pesar de poder tener lugar la hidroxilación normal de la proteína HIF, la proteína anómala pVHL generada a partir de genes mutados (representada con cuadraditos), carece de receptores adecuados de reconocimiento para HIF-hidroxilada, no originándose su poliubiquitinación y posterior degradación, predominado los mecanismos que conducen a agravamiento de la enfermedad y su relación con el cáncer. HIF, factor inducible por hipoxia; pVHL: proteina Von Hippel-Lindau. (Esquema de diseño propio).
Conociendo la importancia de la hidroxilación en el reconocimiento de HIF-1α por pVHL, se inició la búsqueda para identificar las enzimas responsables. La hidroxilación de las prolinas ya había sido definida en la estabilización de las cadenas de procolágeno y existían enormes similitudes con los procesos dependientes de hipoxia, ya que ambas reacciones requerían oxígeno, hierro y ascorbato, y eran inhibidas por el cobalto y los análogos del 2-oxoglutarato. Esto sugirió que la prolil-hidroxilasa que activaba a HIF era un miembro de la familia de la enzima dioxigenasa dependiente de 2-oxoglutrato. Posteriormente, se identificó un grupo de enzimas como los sensores clave de oxígeno basados en el descubrimiento de la proteína egl9 en Caenorhabditis elegans: las proteínas EglN (Egl nine homolog) o PHD (dominio de prolil-hidroxilasa), capaces de inducir la hidroxilación del HIF-1/2α humano(24-26).
Se demostró que EglN1 (también llamada proteína que contiene el dominio de prolil-hidroxilasa-2 [PHD2]) es la hidroxilasa crítica para HIF-α in vivo(27). El requerimiento de 2-oxoglutato para la reacción proporcionó un vínculo adicional entre la hipoxia y el metabolismo. La afinidad tan elevada del EglNs por el oxígeno sugirió que esta molécula era un sensor de oxígeno, una observación que fue confirmada genéticamente. Además, se ha demostrado que las mutaciones inactivadoras en EglN1 o mutaciones activadoras en HIF-2α causan eritrocitosis familiar(28). También, en poblaciones que viven a grandes altitudes, la existencia de polimorfismos de un solo nucleótido o las mutaciones sin-sentido en EglN1 permiten la adaptación a gran altitud(8).
Como colofón a este homenaje, señalaremos que las relaciones, aparentemente sencillas, destacadas por la Fundación Nobel tienen en la actualidad importantes implicaciones farmacológicas no sólo bajo el punto de vista del cáncer, sino de otras afecciones como la anemia y las enfermedades cardiovasculares y pulmonares. En el ámbito del cáncer, mientras que los tumores asociados con VHL y los carcinomas renales muestran un notable estado pseudohipóxico y una dependencia vascular debido a la estabilización constitutiva de HIF-2α, en muchos otros tipos de tumores uno o ambos factores HIF están estabilizados o regulados, lo que con frecuencia puede ser un potente biomarcador de tumores agresivos. Es importante destacar que las pequeñas moléculas que se dirigen a las proteínas HIF están emergiendo como terapias contra el cáncer(29). Respecto a los efectos mediados por la hipoxia señalar que puede originar un cúmulo de especies reactivas de oxígeno e induciendo daño, particularmente en nichos con importantes implicaciones como la médula ósea, pudiendo afectar la formación de células hemáticas y elementos formes sanguíneos.
Referencias
1. Sánchez-Muniz FJ, Culebras JM, Vicente-Vacas L. Rendimos homenaje a Alfred Nobel y a los galardonados con el Premio de Fisiología y Medicina de 2019. JONNPR. 2020;5(3):221-30. DOI: 10.19230/jonnpr.3451
2. Premio Nobel de Medicina 2019: Página Oficial. https://www.nobelprizemedicine.org/the-nobel-prize-in-physiology-or-medicine-2019/
3. Moslehi J, Rathmell WK. The 2019 Nobel Prize honors fundamental discoveries in hypoxia response. J Clin Invest. 2019 Nov 25. pii: 134813. doi: 10.1172/JCI134813.
4. Wang GL, Semenza GL. Purification and characterization of hypoxia-inducible factor 1. J Biol Chem. 1995;270(3):1230–1237.
5. Wagner Grau P. El factor HIF-1 inducido por la hipoxia y la sensibilidad al oxígeno. Rol del hierro intracellular. Acta Med. Per. 2011; 28(3):1-7.
6. Hogenesch JB, Chan WK, Jackiw VH, Brown RC, Gu YZ, Pray-Grant M, Perdew GH, Bradfield CA. Characterization of a subset of the basic helix-loop-helix-PAS superfamily that interacts with components of the dioxin signaling pathway. J Biol Chem. 1997; 272 (13):8581-8593.
7. Hanaoka M; Droma Y; Basnyat B; Ito M; Kobayashi N; Katsuyama Y; Kubo K; Ota M. Genetic Variants in EPAS1 contribute to adaptation to high-altitude hypoxia in Sherpas. PLos One. 2012; 7(12): e50566.
8. Simonson TS, Yang Y, Huff CD, Yun H, Qin G, Witherspoon DJ, Bai Z, Lorenzo FR, Xing J, Jorde LB, Prchal JT, Ge R. Genetic evidence for high-altitude adaptation in Tibet. Science. 2010; 329(5987):72–75.
9. https://es.wikipedia.org/wiki/EPAS1 visitada en 30 de noviembre de 2019.
10. Hogenesch JB; Gu YZ; Jain S; Bradfield CA. The basic-helix-loop-helix-PAS orphan MOP3 forms transcriptionally active complexes with circadian and hypoxia factors. Proc. Natl. Acad. Sci. U.S.A. 1998; 95(10):5474-5479.
11. Forsythe JA, Jiang BH, Iyer NV, Agani F, Leung SW, Koos RD, Semenza GL. Activation of vascular endothelial growth factor gene transcription by hypoxia-inducible factor 1. Mol Cell Biol. 1996; 16(9):4604–4613.
12. Von Hippel-Lindau disease / Genetic and Rare Diseases Information Center (GARD) – an NCATS Program. rarediseases.info.nih.gov. visitada en 30 noviembre de 2019.
13. Maher ER, Glenn GM, Walther M, Maher ER, Neumann HP, Richard S. von Hippel-Lindau disease: a clinical and scientific review. Eur J Hum Genet. 2011; 19(6): 617–623.
14. Friedrich CA. Von Hippel-Lindau syndrome. A pleomorphic condition. Cancer. 1999; 86(11 Suppl): 2478–2482.
15. Kondo K, Kaelin WG. The von Hippel–Lindau tumor suppressor gene. Exp Cell Res. 2001; 264(1):117–125.
16. Nordstrom-O'Brien M1, van der Luijt RB, van Rooijen E, van den Ouweland AM, Majoor-Krakauer DF, Lolkema MP, van Brussel A, Voest EE, Giles RH. Genetic analysis of von Hippel-Lindau disease. Hum. Mutat. 2010; 31(5):521–537.
17. Stebbins CE, Kaelin WG, Pavletich NP. Structure of the VHL-ElonginC-Elongin B complex: implications for VHL tumor suppressor function. Science. 1999; 284(5413):455–461.
18. Maxwell PH, Pugh CW, Ratcliffe PJ. Inducible operation of the erythropoietin 3′ enhancer in multiple cell lines: evidence for a widespread oxygen-sensing mechanism. Proc Natl Acad Sci U S A. 1993; 90(6):2423–2427.
19. Maxwell PH, Wiesener MS, Chang GW, Clifford SC, Vaux EC, Cockman ME, Wykoff CC, Pugh CW, Maher ER, Ratcliffe PJ. The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature. 1999; 399(6733):271–275.
20. Jaakkola P, Mole DR, Tian YM, Wilson MI, Gielbert J, Gaskell SJ, von Kriegsheim A, Hebestreit HF, Mukherji M, Schofield CJ, Maxwell PH, Pugh CW, Ratcliffe PJ. Targeting of HIF-α to the von Hippel-Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science. 2001; 292(5516):468–472.
21. Ivan M, Kondo K, Yang H, Kim W, Valiando J, Ohh M, Salic A, Asara JM, Lane WS, Kaelin WG Jr. HIFalpha targeted for VHL-mediated destruction by proline hydroxylation: implications for O2 sensing. Science. 2001; 292(5516):464–468.
22. Kaelin WG. Von Hippel-Lindau disease. Annu Rev Pathol. 2007; 2:145-173.
23. Bader HL; Hsu T. Systemic VHL gene functions and the VHL disease. FEBS Letters. 2012; 586(11):1562–1569.
24. Epstein AC, Gleadle JM, McNeill LA, Hewitson KS, O'Rourke J, Mole DR, Mukherji M, Metzen E, Wilson MI, Dhanda A, Tian YM, Masson N, Hamilton DL, Jaakkola P, Barstead R, Hodgkin J, Maxwell PH, Pugh CW, Schofield CJ, Ratcliffe PJ. C. elegans EGL-9 and mammalian homologs define a family of dioxygenases that regulate HIF by prolyl hydroxylation. Cell. 2001; 107(1):43–54.
25. Bruick RK, McKnight SL. A conserved family of prolyl-4-hydroxylases that modify HIF. Science. 2001; 294(5545):1337–1340.
26. Ivan M, Haberberger T, Gervasi DC, Michelson KS, Günzler V, Kondo K, Yang H, Sorokina I, Conaway RC, Conaway JW, Kaelin WG Jr. Biochemical purification and pharmacological inhibition of a mammalian prolyl hydroxylase acting on hypoxia-inducible factor. Proc Natl Acad Sci U S A. 2002; 99(21):13459–13464.
27. Minamishima YA, Moslehi J, Bardeesy N, Cullen D, Bronson RT, Kaelin WG. Somatic inactivation of the PHD2 prolyl hydroxylase causes polycythemia and congestive heart failure. Blood. 2008; 111(6):3236–3244.
28. Percy MJ, Furlow PW, Lucas GS, Li X, Lappin TR, McMullin MF, Lee FS. A gain-of-function mutation in the HIF2A gene in familial erythrocytosis. N Engl J Med. 2008; 358(2):162–168.
29. Courtney KD, Infante JR, Lam ET, Figlin RA, Rini BI, Brugarolas J, Zojwalla NJ, Lowe AM, Wang K, Wallace EM, Josey JA, Choueiri TK. Phase I dose-escalation trial of PT2385, a first-in-class hypoxia-inducible factor-2α antagonist in patients with previously treated advanced clear cell renal cell carcinoma. J Clin Oncol. 2018; 36(9):867–874.